

How Does An Enzyme Increase The Rate Of A Chemical Reaction
A primal task of proteins is to act as enzymes—catalysts that increase the rate of virtually all the chemical reactions within cells. Although RNAs are capable of catalyzing some reactions, most biological reactions are catalyzed by proteins. In the absence of enzymatic catalysis, most biochemical reactions are so slow that they would not occur nether the balmy conditions of temperature and pressure level that are compatible with life. Enzymes advance the rates of such reactions by well over a million-fold, and then reactions that would take years in the absence of catalysis can occur in fractions of seconds if catalyzed by the appropriate enzyme. Cells contain thousands of dissimilar enzymes, and their activities decide which of the many possible chemical reactions actually accept identify within the jail cell.
The Catalytic Activity of Enzymes
Similar all other catalysts, enzymes are characterized by two cardinal properties. First, they increase the rate of chemical reactions without themselves being consumed or permanently altered by the reaction. Second, they increase reaction rates without altering the chemical equilibrium between reactants and products.
These principles of enzymatic catalysis are illustrated in the post-obit example, in which a molecule acted upon by an enzyme (referred to equally a substrate [S]) is converted to a product (P) as the result of the reaction. In the absenteeism of the enzyme, the reaction can exist written as follows:
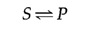
The chemical equilibrium between Due south and P is determined by the laws of thermodynamics (every bit discussed further in the next section of this chapter) and is represented by the ratio of the forward and reverse reaction rates (S→P and P→S, respectively). In the presence of the advisable enzyme, the conversion of Southward to P is accelerated, but the equilibrium between Due south and P is unaltered. Therefore, the enzyme must accelerate both the frontward and opposite reactions equally. The reaction can exist written as follows:
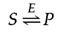
Annotation that the enzyme (Due east) is not altered past the reaction, and then the chemic equilibrium remains unchanged, determined solely by the thermodynamic properties of S and P.
The effect of the enzyme on such a reaction is best illustrated by the energy changes that must occur during the conversion of Southward to P (Figure two.22). The equilibrium of the reaction is determined by the concluding energy states of Due south and P, which are unaffected past enzymatic catalysis. In guild for the reaction to proceed, however, the substrate must first be converted to a college energy state, called the transition state. The free energy required to attain the transition land (the activation free energy) constitutes a barrier to the progress of the reaction, limiting the charge per unit of the reaction. Enzymes (and other catalysts) act by reducing the activation free energy, thereby increasing the rate of reaction. The increased rate is the aforementioned in both the forward and reverse directions, since both must pass through the same transition state.
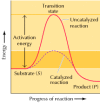
Figure ii.22
Energy diagrams for catalyzed and uncatalyzed reactions. The reaction illustrated is the simple conversion of a substrate S to a product P. Considering the last energy state of P is lower than that of S, the reaction gain from left to right. For the (more...)
The catalytic action of enzymes involves the bounden of their substrates to course an enzyme-substrate complex (ES). The substrate binds to a specific region of the enzyme, called the active site. While bound to the active site, the substrate is converted into the production of the reaction, which is and so released from the enzyme. The enzyme-catalyzed reaction tin thus be written every bit follows:
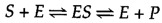
Note that East appears unaltered on both sides of the equation, so the equilibrium is unaffected. However, the enzyme provides a surface upon which the reactions converting S to P can occur more readily. This is a consequence of interactions between the enzyme and substrate that lower the energy of activation and favor formation of the transition state.
Mechanisms of Enzymatic Catalysis
The bounden of a substrate to the active site of an enzyme is a very specific interaction. Active sites are clefts or grooves on the surface of an enzyme, usually composed of amino acids from dissimilar parts of the polypeptide chain that are brought together in the tertiary construction of the folded protein. Substrates initially bind to the active site by noncovalent interactions, including hydrogen bonds, ionic bonds, and hydrophobic interactions. Once a substrate is spring to the agile site of an enzyme, multiple mechanisms can accelerate its conversion to the product of the reaction.
Although the simple example discussed in the previous department involved but a single substrate molecule, almost biochemical reactions involve interactions between 2 or more than different substrates. For example, the formation of a peptide bond involves the joining of two amino acids. For such reactions, the binding of 2 or more substrates to the agile site in the proper position and orientation accelerates the reaction (Figure 2.23). The enzyme provides a template upon which the reactants are brought together and properly oriented to favor the formation of the transition state in which they collaborate.

Effigy 2.23
Enzymatic catalysis of a reaction between ii substrates. The enzyme provides a template upon which the two substrates are brought together in the proper position and orientation to react with each other.
Enzymes accelerate reactions also by altering the conformation of their substrates to approach that of the transition state. The simplest model of enzyme-substrate interaction is the lock-and-key model, in which the substrate fits precisely into the active site (Figure 2.24). In many cases, however, the configurations of both the enzyme and substrate are modified by substrate bounden—a process called induced fit. In such cases the conformation of the substrate is contradistinct so that it more closely resembles that of the transition state. The stress produced by such distortion of the substrate can further facilitate its conversion to the transition state by weakening disquisitional bonds. Moreover, the transition state is stabilized by its tight binding to the enzyme, thereby lowering the required energy of activation.

Effigy 2.24
Models of enzyme-substrate interaction. (A) In the lock-and-key model, the substrate fits precisely into the agile site of the enzyme. (B) In the induced-fit model, substrate binding distorts the conformations of both substrate and enzyme. This baloney (more...)
In addition to bringing multiple substrates together and distorting the conformation of substrates to approach the transition state, many enzymes participate directly in the catalytic process. In such cases, specific amino acid side chains in the agile site may react with the substrate and form bonds with reaction intermediates. The acidic and basic amino acids are often involved in these catalytic mechanisms, as illustrated in the following discussion of chymotrypsin as an example of enzymatic catalysis.
Chymotrypsin is a member of a family unit of enzymes (serine proteases) that digest proteins by catalyzing the hydrolysis of peptide bonds. The reaction can be written as follows:

The unlike members of the serine protease family unit (including chymotrypsin, trypsin, elastase, and thrombin) have distinct substrate specificities; they preferentially cleave peptide bonds adjacent to different amino acids. For instance, whereas chymotrypsin digests bonds adjacent to hydrophobic amino acids, such as tryptophan and phenylalanine, trypsin digests bonds next to basic amino acids, such every bit lysine and arginine. All the serine proteases, however, are similar in structure and employ the same mechanism of catalysis. The active sites of these enzymes contain three critical amino acids—serine, histidine, and aspartate—that drive hydrolysis of the peptide bond. Indeed, these enzymes are called serine proteases because of the central role of the serine balance.
Substrates bind to the serine proteases past insertion of the amino acid adjacent to the cleavage site into a pocket at the agile site of the enzyme (Figure 2.25). The nature of this pocket determines the substrate specificity of the different members of the serine protease family. For example, the binding pocket of chymotrypsin contains hydrophobic amino acids that collaborate with the hydrophobic side chains of its preferred substrates. In dissimilarity, the binding pocket of trypsin contains a negatively charged acidic amino acid (aspartate), which is able to form an ionic bail with the lysine or arginine residues of its substrates.
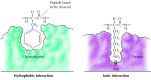
Figure ii.25
Substrate bounden by serine proteases. The amino acid adjacent to the peptide bail to be cleaved is inserted into a pocket at the active site of the enzyme. In chymotrypsin, the pocket binds hydrophobic amino acids; the binding pocket of trypsin contains (more...)
Substrate binding positions the peptide bail to be cleaved adjacent to the active site serine (Figure 2.26). The proton of this serine is then transferred to the active site histidine. The conformation of the active site favors this proton transfer because the histidine interacts with the negatively charged aspartate residue. The serine reacts with the substrate, forming a tetrahedral transition state. The peptide bond is so cleaved, and the C-terminal portion of the substrate is released from the enzyme. Still, the North-terminal peptide remains bound to serine. This situation is resolved when a water molecule (the second substrate) enters the active site and reverses the preceding reactions. The proton of the h2o molecule is transferred to histidine, and its hydroxyl group is transferred to the peptide, forming a 2d tetrahedral transition land. The proton is and then transferred from histidine back to serine, and the peptide is released from the enzyme, completing the reaction.

Effigy 2.26
Catalytic machinery of chymotrypsin. 3 amino acids at the active site (Ser-195, His-57, and Asp-102) play disquisitional roles in catalysis.
This case illustrates several features of enzymatic catalysis; the specificity of enzyme-substrate interactions, the positioning of dissimilar substrate molecules in the active site, and the interest of active-site residues in the formation and stabilization of the transition state. Although the thousands of enzymes in cells catalyze many dissimilar types of chemic reactions, the same basic principles apply to their functioning.
Coenzymes
In addition to bounden their substrates, the active sites of many enzymes bind other pocket-sized molecules that participate in catalysis. Prosthetic groups are small molecules bound to proteins in which they play critical functional roles. For example, the oxygen carried by myoglobin and hemoglobin is bound to heme, a prosthetic grouping of these proteins. In many cases metal ions (such as zinc or iron) are leap to enzymes and play central roles in the catalytic procedure. In addition, diverse low-molecular-weight organic molecules participate in specific types of enzymatic reactions. These molecules are called coenzymes because they piece of work together with enzymes to heighten reaction rates. In contrast to substrates, coenzymes are not irreversibly altered by the reactions in which they are involved. Rather, they are recycled and can participate in multiple enzymatic reactions.
Coenzymes serve as carriers of several types of chemical groups. A prominent example of a coenzyme is nicotinamide adenine dinucleotide (NAD +), which functions as a carrier of electrons in oxidation-reduction reactions (Effigy two.27). NAD+ tin can accept a hydrogen ion (H+) and two electrons (e-) from one substrate, forming NADH. NADH tin can then donate these electrons to a 2nd substrate, re-forming NAD+. Thus, NAD+ transfers electrons from the get-go substrate (which becomes oxidized) to the 2d (which becomes reduced).

Figure 2.27
Office of NAD+ in oxidation-reduction reactions. (A) Nicotinamide adenine dinucleotide (NAD+) acts every bit a carrier of electrons in oxidation-reduction reactions past accepting electrons (east-) to form NADH. (B) For case, NAD+ can accept electrons from one substrate (more...)
Several other coenzymes too human activity as electron carriers, and still others are involved in the transfer of a variety of additional chemic groups (e.chiliad., carboxyl groups and acyl groups; Tabular array 2.1). The same coenzymes function together with a diversity of unlike enzymes to catalyze the transfer of specific chemical groups between a wide range of substrates. Many coenzymes are closely related to vitamins, which contribute role or all of the structure of the coenzyme. Vitamins are not required by bacteria such as Eastward. coli but are necessary components of the diets of human being and other college animals, which take lost the ability to synthesize these compounds.
Regulation of Enzyme Action
An important feature of virtually enzymes is that their activities are non constant just instead can be modulated. That is, the activities of enzymes can be regulated so that they function appropriately to meet the varied physiological needs that may ascend during the life of the cell.
I common type of enzyme regulation is feedback inhibition, in which the product of a metabolic pathway inhibits the activeness of an enzyme involved in its synthesis. For example, the amino acrid isoleucine is synthesized by a series of reactions starting from the amino acrid threonine (Figure 2.28). The first step in the pathway is catalyzed by the enzyme threonine deaminase, which is inhibited by isoleucine, the end product of the pathway. Thus, an acceptable amount of isoleucine in the cell inhibits threonine deaminase, blocking further synthesis of isoleucine. If the concentration of isoleucine decreases, feedback inhibition is relieved, threonine deaminase is no longer inhibited, and boosted isoleucine is synthesized. By so regulating the activity of threonine deaminase, the prison cell synthesizes the necessary amount of isoleucine just avoids wasting energy on the synthesis of more isoleucine than is needed.

Figure 2.28
Feedback inhibition. The first step in the conversion of threonine to iso-leucine is catalyzed by the enzyme threonine deaminase. The activity of this enzyme is inhibited by isoleucine, the end product of the pathway.
Feedback inhibition is one example of allosteric regulation, in which enzyme activity is controlled by the binding of pocket-size molecules to regulatory sites on the enzyme (Effigy 2.29). The term "allosteric regulation" derives from the fact that the regulatory molecules bind not to the catalytic site, but to a singled-out site on the protein (allo= "other" and steric= "site"). Bounden of the regulatory molecule changes the conformation of the protein, which in turn alters the shape of the agile site and the catalytic activity of the enzyme. In the instance of threonine deaminase, binding of the regulatory molecule (isoleucine) inhibits enzymatic activity. In other cases regulatory molecules serve as activators, stimulating rather than inhibiting their target enzymes.
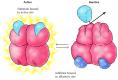
Figure two.29
Allosteric regulation. In this example, enzyme action is inhibited by the binding of a regulatory molecule to an allosteric site. In the absence of inhibitor, the substrate binds to the active site of the enzyme and the reaction proceeds. The binding (more...)
The activities of enzymes can also be regulated by their interactions with other proteins and past covalent modifications, such as the add-on of phosphate groups to serine, threonine, or tyrosine residues. Phosphorylation is a particularly common mechanism for regulating enzyme action; the addition of phosphate groups either stimulates or inhibits the activities of many dissimilar enzymes (Effigy 2.xxx). For example, musculus cells respond to epinephrine (adrenaline) past breaking down glycogen into glucose, thereby providing a source of energy for increased muscular activity. The breakdown of glycogen is catalyzed by the enzyme glycogen phosphorylase, which is activated past phosphorylation in response to the bounden of epinephrine to a receptor on the surface of the muscle cell. Protein phosphorylation plays a central role in controlling non but metabolic reactions but too many other cellular functions, including cell growth and differentiation.

Figure 2.30
Protein phosphorylation. Some enzymes are regulated past the addition of phosphate groups to the side-concatenation OH groups of serine (equally shown here), threonine, or tyrosine residues. For example, the enzyme glycogen phosphorylase, which catalyzes the conversion (more...)

How Does An Enzyme Increase The Rate Of A Chemical Reaction,
Source: https://www.ncbi.nlm.nih.gov/books/NBK9921/
Posted by: sheltonbarl1969.blogspot.com
0 Response to "How Does An Enzyme Increase The Rate Of A Chemical Reaction"
Post a Comment